Віруси - це не просто надокучливі збудники хвороб. Це давні та загадкові біологічні агенти, історія яких сягає глибин еволюції. Чи були вони колись вільноживучими організмами? Чи, можливо, вони існували ще до появи клітин?
Які існують думки?
Серед вірусологів точаться запеклі дебати з цього питання. Було сформульовано три основні гіпотези: 1) прогресивна гіпотеза, або гіпотеза виходу, стверджує, що віруси виникли з генетичних елементів, які набули здатності переміщатися між клітинами; 2) регресивна гіпотеза, або гіпотеза редукції, стверджує, що віруси є залишками клітинних організмів; 3) гіпотеза "віруси першими" стверджує, що віруси існували до або еволюціонували разом зі своїми нинішніми клітинними хазяями.
Прогресивна гіпотеза
Згідно з цією гіпотезою, ретровіруси могли виникнути, коли ретротранспозони набули здатності залишати одну клітину та інфікувати іншу. Цьому сприяла поява у них структурних білків, які дозволили формувати вірусні частинки.
Ретровіруси мають одноланцюговий РНК-геном. Коли вірус проникає в клітину-хазяїна, вірусний фермент, зворотна транскриптаза, перетворює цю одноланцюгову РНК на дволанцюгову ДНК. Ця вірусна ДНК потім мігрує до ядра клітини-хазяїна. Інший вірусний фермент, інтеграза, вбудовує новоутворену вірусну ДНК у геном клітини-хазяїна. Потім вірусні гени можуть транскрибуватися та транслюватися. РНК-полімераза клітини-хазяїна може виробляти нові копії одноланцюгового РНК-геному вірусу. Дочірні віруси збираються та виходять з клітини, щоб розпочати процес знову.
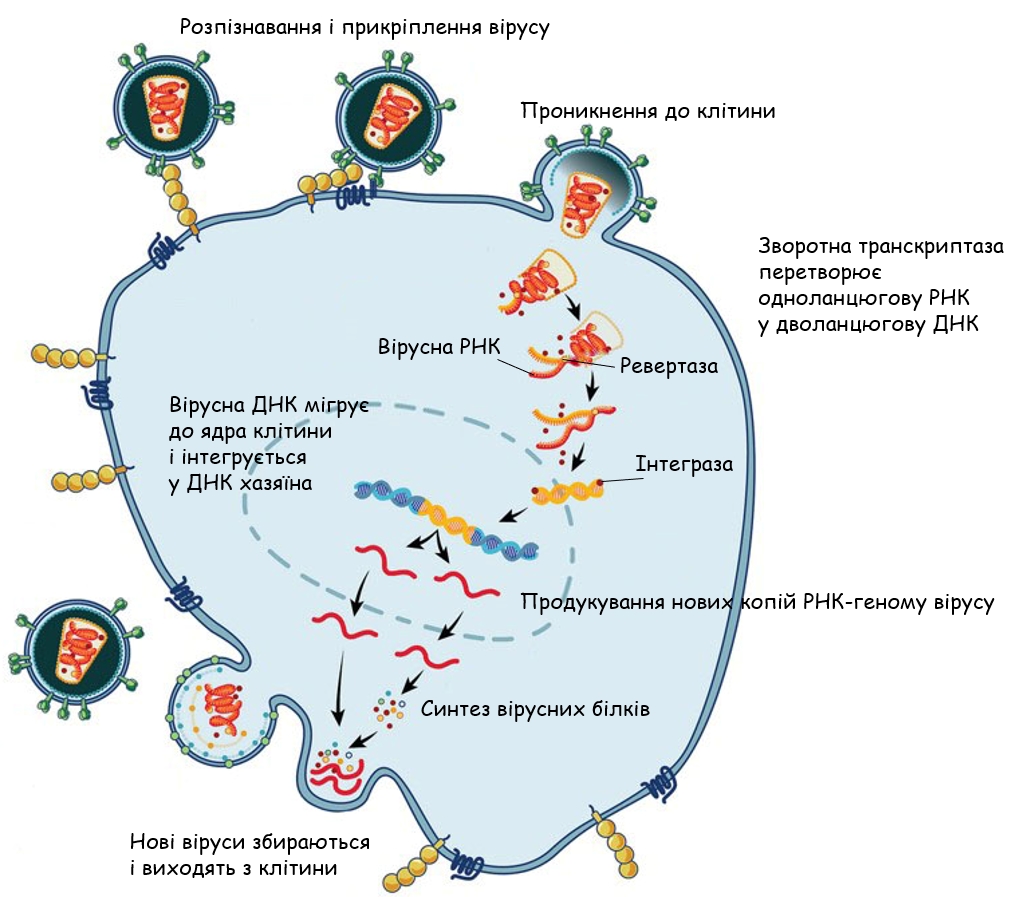
Цей процес дуже схожий на переміщення важливого, хоча й дещо незвичайного, компонента більшості еукаріотичних геномів: ретротранспозонів. Ці мобільні генетичні елементи складають вражаючі 42% геному людини і можуть переміщатися в межах геному за допомогою РНК-посередника. Як і ретровіруси, певні класи ретротранспозонів, вірусоподібні ретротранспозони, кодують зворотну транскриптазу і, часто, інтегразу. За допомогою цих ферментів ці елементи можуть транскрибуватися в РНК, зворотно транскрибуватися в ДНК, а потім інтегруватися в нове місце в геномі. Можна припустити, що придбання кількох структурних білків дозволило б елементу вийти з клітини та проникнути в нову клітину, перетворившись таким чином на інфекційний агент. Дійсно, генетичні структури ретровірусів та вірусоподібних ретротранспозонів виявляють разючу схожість.
Деградація
На відміну від описаного вище прогресивного процесу, віруси могли виникнути в результаті регресивного, або редукційного, процесу. Мікробіологи загалом погоджуються, що певні бактерії, які є облігатними внутрішньоклітинними паразитами, такі як види Chlamydia та Rickettsia, еволюціонували від вільноживучих предків. Дійсно, геномні дослідження показують, що мітохондрії еукаріотичних клітин та Rickettsia prowazekii можуть мати спільного вільноживучого предка. Звідси випливає, що існуючі віруси могли еволюціонувати з більш складних, можливо, вільноживучих організмів, які з часом втратили генетичну інформацію, оскільки вони перейшли до паразитичного способу реплікації.
Віруси однієї конкретної групи, великі ядерно-цитоплазматичні ДНК-вмісні віруси (NCLDV), найкраще ілюструють цю гіпотезу. Ці віруси, до яких належать вірус віспи та нещодавно відкритий гігант серед усіх вірусів, мімівірус, набагато більші за більшість вірусів. Наприклад, типовий вірус віспи у формі цеглини може мати ширину 200 нм і довжину 300 нм.
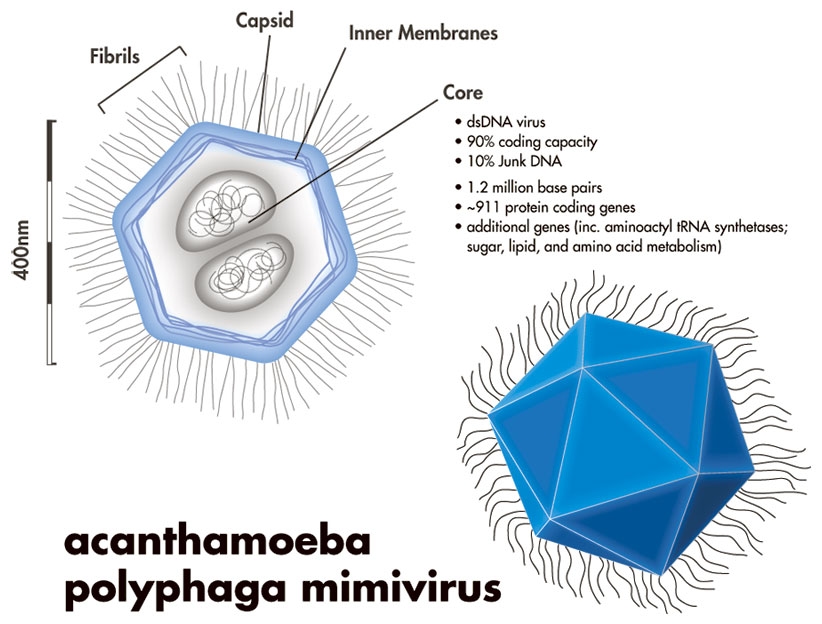
Мімівірус, який приблизно вдвічі більший, має загальний діаметр близько 750 нм. І навпаки, сферичні частинки вірусу грипу можуть мати діаметр лише 80 нм, а частинки поліовірусу - лише 30 нм, що приблизно в 10 000 разів менше за крупинку солі. NCLDV також мають великі геноми. Знову ж таки, геноми вірусу віспи часто наближаються до 200 000 пар основ, а мімівірус має геном з 1,2 мільйона пар основ; тоді як поліовірус має геном лише з 7500 нуклеотидів. На додаток до своїх великих розмірів, NCLDV демонструють більшу складність, ніж інші віруси, і менше залежать від свого хазяїна для реплікації, ніж інші віруси. Наприклад, частинки вірусу віспи містять велику кількість вірусних ферментів та споріднених факторів, які дозволяють вірусу виробляти функціональну матричну РНК у цитоплазмі клітини-хазяїна.
Через розмір і складність NCLDV деякі вірусологи висунули гіпотезу, що ці віруси можуть бути нащадками більш складних предків. На думку прихильників цієї гіпотези, автономні організми спочатку розвинули симбіотичні відносини. З часом ці відносини стали паразитичними, оскільки один організм ставав все більш залежним від іншого. У міру того, як колись вільноживучий паразит ставав все більш залежним від хазяїна, він втрачав раніше життєво важливі гени. Зрештою, він втратив здатність до самостійної реплікації, перетворившись на облігатного внутрішньоклітинного паразита, тобто вірус. Аналіз гігантського мімівірусу може підтвердити цю гіпотезу. Цей вірус містить відносно великий репертуар передбачуваних генів, пов'язаних з трансляцією, - генів, які можуть бути залишками раніше повної системи трансляції. Цікаво, що мімівірус не сильно відрізняється від паразитичних бактерій, таких як Rickettsia prowazekii.
Гіпотеза "віруси першими"
А що, як віруси існували першими? Нещодавно кілька дослідників висунули припущення, що віруси, можливо, були першими реплікуючими одиницями. Кунін і Мартін постулювали, що віруси існували в доклітинному світі як самовідтворювані одиниці. З часом ці одиниці, на їхню думку, ставали все більш організованими та складними. Зрештою, еволюціонували ферменти для синтезу мембран і клітинних стінок, що призвело до утворення клітин. Таким чином, віруси, можливо, існували до бактерій, архей або еукаріотів.
Більшість біологів зараз погоджуються, що перші реплікуючі молекули складалися з РНК, а не з ДНК. Ми також знаємо, що деякі молекули РНК, рибозими, мають ферментативні властивості; вони можуть каталізувати хімічні реакції. Можливо, прості реплікуючі молекули РНК, що існували до утворення першої клітини, розвинули здатність інфікувати перші клітини. Чи можуть сьогоднішні одноланцюгові РНК-вмісні віруси бути нащадками цих доклітинних молекул РНК?
Інші стверджують, що попередники сьогоднішніх NCLDV призвели до появи еукаріотичних клітин. Вільярреал і ДеФіліппіс та Белл описали моделі, що пояснюють цю гіпотезу. Можливо, обидві групи припускають, що нинішнє ядро в еукаріотичних клітинах виникло в результаті ендосимбіотичної події, коли складний, оточений оболонкою ДНК-вмісний вірус став постійним мешканцем еукаріотичної клітини, що формувалася. Такими чином, віруси еволюціонували з фундаментальних реплікативних молекул, які утворилися в «первісному супі», коли планета почала охолоджуватися. Ці молекули також призвели до еволюції клітинних організмів - вірусних господарів - або паралельно, або на пізній стадії еволюції. Ранні віруси могли існувати як вільно плаваючі шматочки нуклеїнових кислот, які стали більш організованими та складнішими.
Жодна гіпотеза не може бути повністю вірною
Звідки взялися віруси - непросте питання. Можна досить переконливо стверджувати, що певні віруси, такі як ретровіруси, виникли в результаті прогресивного процесу. Мобільні генетичні елементи набули здатності переміщатися між клітинами, перетворюючись на інфекційні агенти. Також можна стверджувати, що великі ДНК-вмісні віруси виникли в результаті регресивного процесу, в ході якого колись незалежні об'єкти з часом втратили ключові гени та перейшли до паразитичної стратегії реплікації. Нарешті, ідея про те, що віруси дали початок життю в тому вигляді, в якому ми його знаємо, відкриває дуже цікаві можливості. Можливо, сьогоднішні віруси виникли кілька разів, за допомогою різних механізмів. Можливо, всі віруси виникли за допомогою механізму, який ще належить відкрити. Сьогоднішні фундаментальні дослідження в таких галузях, як мікробіологія, геноміка та структурна біологія, можуть дати нам відповіді на це фундаментальне питання.
Словничок:
Ретротранспозони: Мобільні генетичні елементи, які копіюють себе та вставляють копії в інші ділянки геному.
Зворотна транскриптаза: Фермент, який каталізує синтез ДНК на матриці РНК.
Інтеграза: Фермент, який вбудовує вірусну ДНК у геном клітини-хазяїна.
Структурні білки: Білки, які формують оболонку вірусу та забезпечують його стабільність поза клітиною.
Капсид: Білкова оболонка, яка оточує генетичний матеріал вірусу.
Рибосоми: Клітинні органели, відповідальні за синтез білків.
АТФ (аденозинтрифосфат): Основне джерело енергії для клітин.
Трансляція: Процес синтезу білків з матричної РНК.
Облігатні внутрішньоклітинні паразити: Організми, які можуть розмножуватися лише всередині клітин інших організмів.
Література:
https://www.sciencedirect.com/science/article/abs/pii/S0168170206000293
https://pmc.ncbi.nlm.nih.gov/articles/PMC7173561/
https://www.nature.com/articles/s41579-019-0205-6
https://www.sciencedirect.com/science/article/abs/pii/S0923250809001065
https://pubmed.ncbi.nlm.nih.gov/11523012/
https://pubmed.ncbi.nlm.nih.gov/16223546/
Сподобалася стаття? Підтримай автора донатом на каву з вірусом (або без, якщо боїшся)! Посилання на банку в закріплених повідомленнях. А ще не забудь підписатися, щоб не пропустити нові захопливі матеріали!

